What Is Dissolved Carbon Dioxide?
We usually associate carbon dioxide (CO₂) with its gaseous state, which is the primary form found on Earth under standard temperature and pressure (STP). By contrast, on Mars, CO₂ exists mainly as dry ice or in liquid form due to the planet’s colder temperatures and lower atmospheric pressure.
In the gas phase, molecules are widely spaced, giving gases their ability to expand uniformly and exert pressure. For example, water vapor (H₂O) is a gas. In the liquid phase, molecules are closer together and held by weak intermolecular forces. Liquid H₂O, for instance, forms a network of hydrogen bonds that contributes to its hydrostatic pressure — the pressure a fluid exerts at rest under gravity.
When CO₂ dissolves in water, it forms dissolved CO₂ (dCO₂). This occurs when individual CO₂ molecules become surrounded by and weakly bonded to H₂O molecules via Van der Waals forces. Ruminal dCO₂ is therefore a liquid, with a distinct infrared signature that lies between liquid H₂O and gaseous CO₂. Unlike gas, ruminal dCO₂ does not generate pressure; it is a liquid, that is denser than H₂O and tends to sink.
Dissolved CO₂ plays critical roles in biology. It is essential to the ruminal buffering system, to photosynthesis in stomata, to gas exchange in the lungs, to blood and cellular buffering, and to nutrient absorption in the gastrointestinal tract. In the rumen, dCO₂ is produced during microbial fermentation and by the ruminal epithelium as part of the proton-buffering process associated with SCFA absorption (Figure below).
This ruminal model differs from other aqueous environments, such as the ocean, where the main source of CO₂ is atmospheric pressure. In those systems, the partial pressure of CO₂ gas controls dCO₂ concentrations via Henry’s law and the Bunsen coefficient. By contrast, ruminal fluid is a non-ideal solution, meaning Henry’s law does not apply; ruminal dCO₂ concentrations are not linearly related to the CO₂ partial pressure in the gas cap.
As a result, using the Bunsen coefficient to calculate ruminal dCO₂ concentrations can lead to significant underestimation. This is illustrated by the following example from Moate et al. (1997).
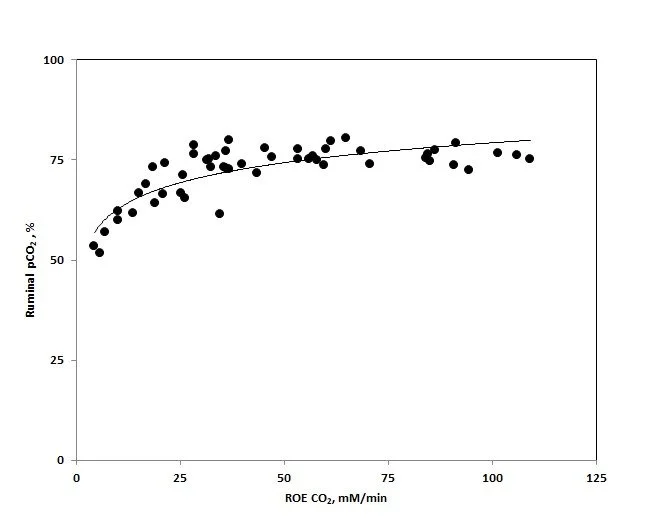
Figure 1 illustrates the relationship between the partial pressure of CO₂ (pCO₂) in the ruminal gas cap (Y-axis) and the rate of entry (ROE) of CO₂ (X-axis). ROE is an indirect measure of dCO₂ concentrations. As shown, pCO₂ remains relatively stable between 50–80%, whereas ROE can range from 0 to 130 mM. This demonstrates the non-ideal behavior of ruminal fluid, where Henry’s law cannot be applied to accurately predict dissolved CO₂ concentrations.
The ruminal buffering system depends on the rapid release of CO₂ gas from the liquid phase. When this effervescence is impaired, dCO₂ accumulates — a condition known as CO₂ holdup. This phenomenon occurs when the physicochemical properties of ruminal fluid are disturbed, preventing efficient gas release.
Accumulation of dCO₂ has multiple physiological and nutritional consequences, several of which have been highlighted in my previous publications. The key challenge remains:
What is the most effective method to measure this elusive but critical molecule in vivo?

The rumen buffer system dominated by CO2 species and water